



















清华大学生命学院吝易、杨雪瑞团队揭示相分离调控节律性蛋白翻译的分子机制
2024/04/09
 1004
1004“朝搴阰之木兰兮,夕揽洲之宿莽”“晨兴理荒秽,带月荷锄归”,地球自转造就日月流转,昼夜更替。为适应这种规律,在宏观层面上,人类活动在时间和空间上随昼夜变化;而在微观层面上,细胞中的众多生物过程也随着昼夜节律在时间和空间上被严格调控,通过精妙地协调多种生物过程的振荡形成无形的生物钟。
6月26日,清华大学生命科学学院、清华-IDG/麦戈文脑科学研究院、清华-北大生命科学联合中心吝易课题组与清华大学生命科学学院杨雪瑞课题组合作在知名刊物《细胞》(Cell)发表了题为”区室化振荡翻译调节昼夜节律时钟”(Circadian Clocks are Modulated by Compartmentalized Oscillating Translation)的研究论文,揭示了细胞中利用相分离协调蛋白质翻译的精细时空调控,从而维持昼夜节律周期的分子机制。
研究人员首先聚焦于细胞内全局性蛋白节律性翻译的分子调控机制。在细胞节律同步化后,研究人员发现细胞内蛋白翻译存在节律性振荡且翻译高峰出现在节律时间CT(circadian time)12-16h。而小鼠在自然昼夜节律下,包括脑及肝在内的组织层面的蛋白翻译也存在节律性振荡。为了进一步研究调控此节律性翻译的分子机制,研究人员分析了不同节律时间的全蛋白组,并初步找到了与蛋白翻译节律振荡模式类似的RNA结合蛋白ATXN2及其同源蛋白ATXN2L为潜在调控因子。有趣的是,ATXN2和ATXN2L单敲除的细胞节律相位向相反方向变化,显示二者在不同时间点发挥功能并协同调控蛋白翻译(图1)。为了进一步探究ATXN2/2L在翻译中的调控作用,研究人员使用了多核糖体图谱分析技术(ribosome profiling),结果显示,翻译高峰时间CT12与翻译低谷时间CT4相比,野生型细胞中大量基因出现明显的翻译激活,而这群基因中一大部分的翻译在ATXN2/2L双敲除的细胞中无法被激活,表明ATXN2和ATXN2L为节律性翻译的主调控分子。
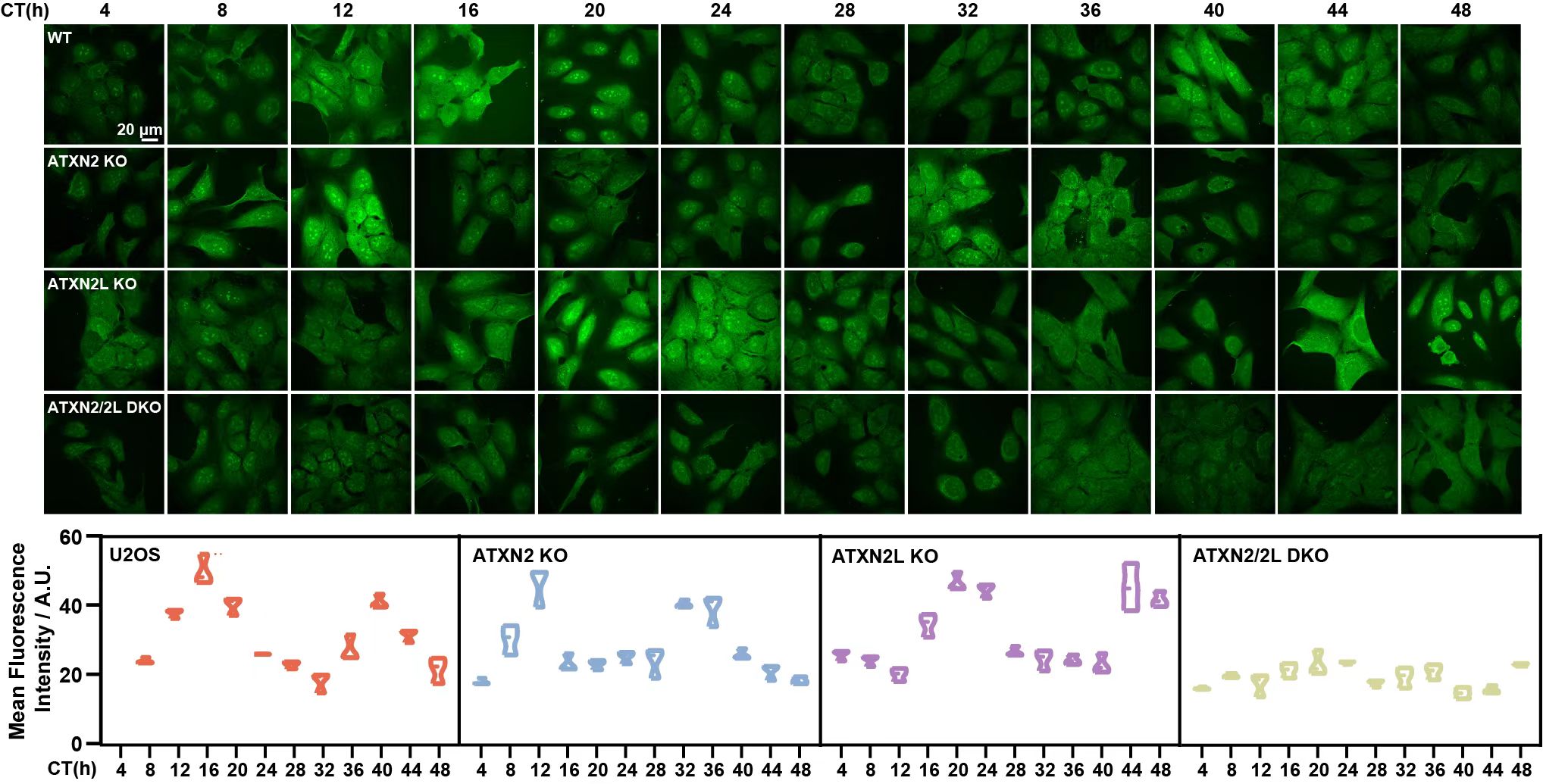
图1.野生型细胞、ATXN2/2L单敲除及双敲除细胞的整体翻译激活情况
通过对细胞和组织的染色观察,研究人员发现ATXN2/2L在哺乳动物脑内中枢生物钟所在的视交叉上核(Suprachiasmatic Nucleus,SCN)以及体外培养的细胞中都会形成蛋白凝聚体(图2)。这些凝聚体位于核周且其大小及数量随昼夜节律振荡。研究人员通过Cryo-mill-IP的方法分离并分析了不同时间点ATXN2凝聚体中的蛋白组分,结果显示,ATXN2凝聚体依时序招募并激活一系列的生化反应,包括在CT12时招募RNA加工相关蛋白,在CT16时大量募集翻译起始因子和核糖体蛋白。
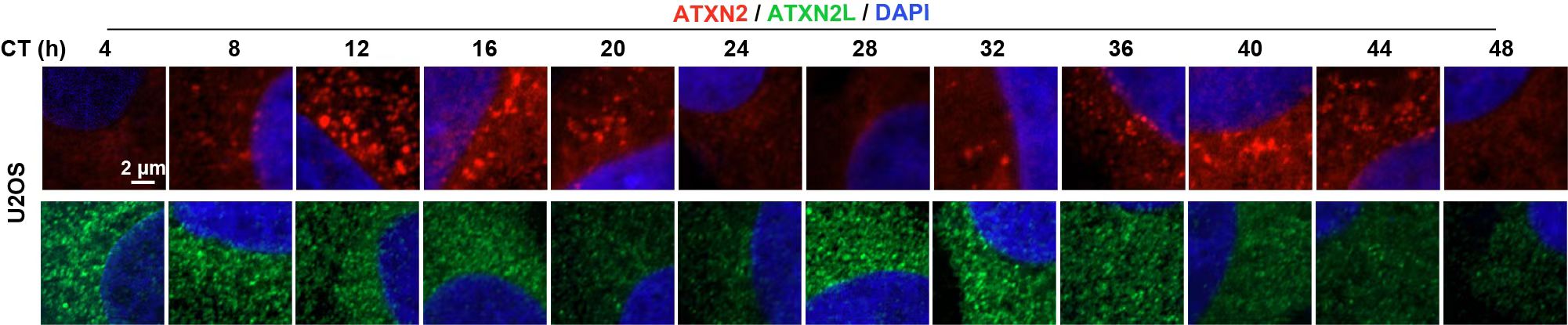
图2.ATXN2/2L凝聚体大小及数量在U2OS细胞中随昼夜节律振荡
之前的研究表明,生物钟调控由若干分子环路组成,其中包括一组时钟基因(clock genes)构成的转录翻译负反馈回路(Transcription-Translation Feedback Loop,TTFL)。哺乳动物的TTFL中,转录激活因子CLOCK和BMAL1激活PER和CRY的转录,而Per和Cry蛋白异二聚体转移回细胞核,结合CLOCK-BMAL1并停止转录激活。后续一系列实验发现ATXN2/2L通过相分离参与到节律蛋白的翻译调控中。ATXN2作为RNA结合蛋白,其特异性地与RNA中的AUUUU/A motif结合,研究人员发现若干节律核心基因,例如PER2RNA的3'非翻译区(3'-UTR)富含该段序列。借助ATXN2体外液-液相分离(LLPS)的特性,研究人员证实ATXN2凝聚体能特异性招募富含该motif的节律分子RNA,而其他对照相分离液滴不具备该特性。在SCN中,同样可以观察到ATXN2凝聚体与PER2RNA的共定位(图3)。
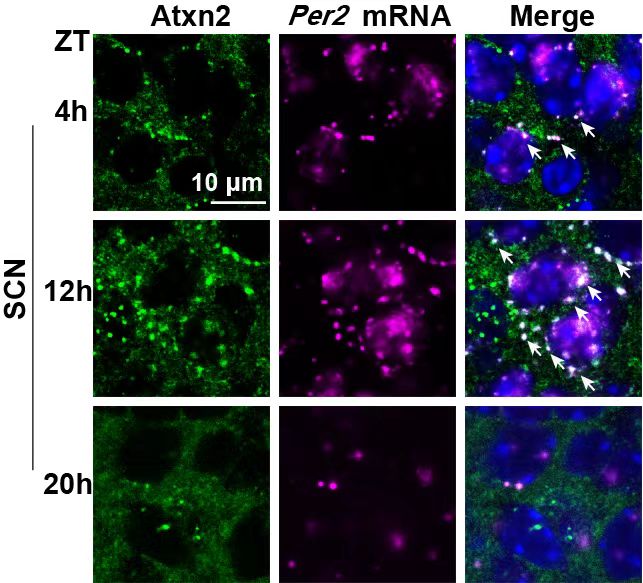
图3. ATXN2凝聚体在SCN中与PER2mRNA共定位
为了建立相分离和翻译调控的因果关系,研究人员利用体外翻译系统证明ATXN2相分离能促进PER2RNA的翻译,并且在细胞内可以直接观察到PER2蛋白在ATXN2凝聚体中合成的过程(图4)。
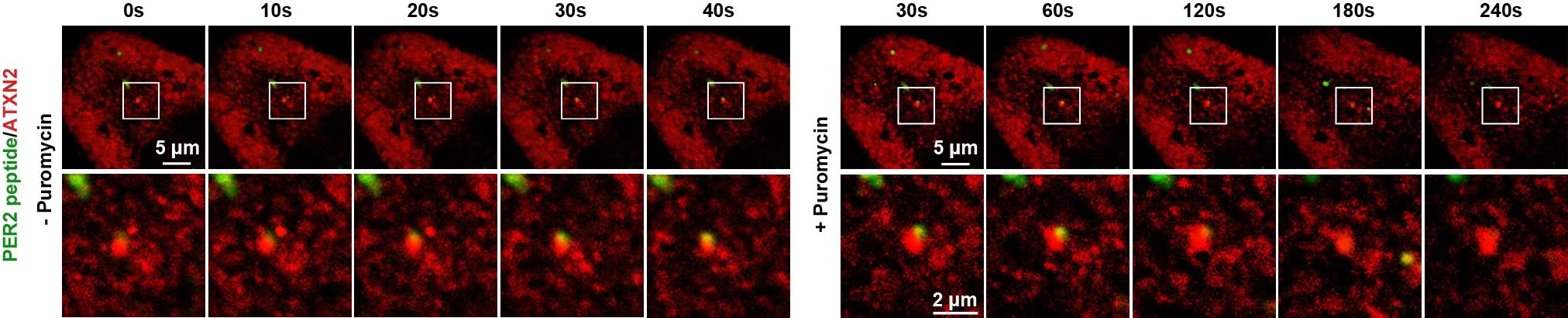
图4.利用SunTag技术观察PER2蛋白在ATXN2凝聚体中合成
通过对ATXN2/2L双敲除细胞进行回补实验,研究人员证实ATXN2野生型可以回补细胞全局翻译以及节律基因的振荡表型,而相分离突变体无法回补相关节律表型。
最后,研究人员将所发现机制在动物层面进行了验证。他们对野生型和ATXN2/2L基因敲除小鼠进行了运动节律行为检测,并比较了节律参数的变化。实验发现,在SCN中条件性敲除ATXN2或2L后,小鼠昼夜节律的周期长度发生相反方向的变化,而当两个基因同时敲除后,小鼠节律的稳定性显著下降。
综上,此项研究发现ATXN2和ATXN2L在细胞内发生相分离,且其相分离随昼夜节律振荡,阐释了其相分离形成的液滴随节律振荡并依序招募和富集一系列RNA相关的生物过程,并在振荡高峰期募集核糖体和特异RNA,促进关键节律蛋白的翻译。这个在翻译上的加速环节在整个TTFL中起到了稳定和精确调控节律周期的作用(图5)。
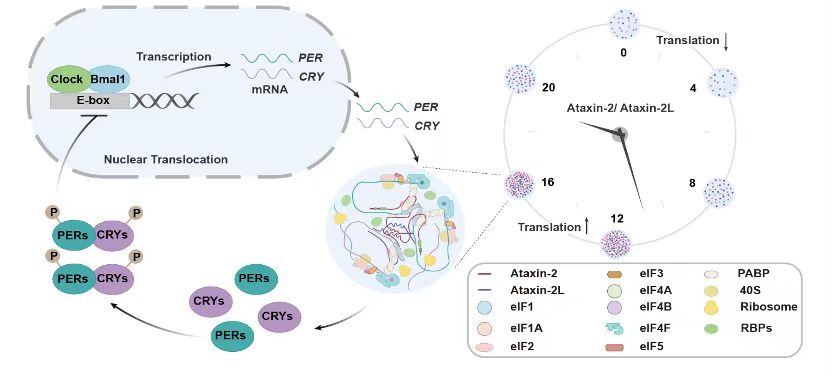
图5.ATXN2/2L相分离调控节律性蛋白翻译的分子机制模型图
相分离作为一种重要的物理化学机制,参与调控了细胞内的众多生理过程。生物大分子发生相分离并形成无膜细胞器(membrane-less organelles, MLOs),使细胞内生化反应区室化,从而对各种生理活动实现精细的时空调控。本文揭示了相分离调节昼夜节律中翻译的功能。之前关于转录调控的研究表明,不同蛋白凝聚体在转录过程的不同阶段,包括起始、延伸和终止发挥功能。本研究中揭示的相分离对于翻译起始阶段的促进进一步支持了细胞内信息液相流动(liquid information flow)的模型。
此外,之前大多数相分离的研究系统是在生理稳态或单向转变(如压力以及细胞分化条件)的细胞环境下进行的。本文中随昼夜节律振荡的相分离系统为未来研究相分离对各种细胞活动的时空调节提供全新的视角。此外,ATXN2是一个与多种神经退行性疾病密切相关的蛋白,因此本研究也对未来的疾病研究具有指导意义,未来相关疾病的治疗应根据文中揭示的ATXN2凝聚体的节律振荡特性仔细考虑诊断和治疗干预的时机。
清华大学生命科学学院吝易实验室2019级博士研究生庄妍榕为论文第一作者,清华大学生命科学学院2018级博士研究生李志原、2021级博士研究生熊诗玥、2018级博士研究生孙楚杰为本文的共同第一作者。清华大学生命学院吝易与杨雪瑞研究员为共同通讯作者。研究得到国家自然科学基金委员会、科技部、膜生物学国家重点实验室、生物信息学教育部重点实验室的大力支持。
文章来源清华大学新闻,分享只为学术交流,如涉及侵权问题请联系我们,我们将及时修改或删除。







-
2026年第五届机器学习、云计算与智 26

-
2026年第二届计算机视觉与机器学习 627
-
2026年6月优质国际学术会议推荐 1157
-
2026年智慧教育与数据挖掘国际学术 813
-
2026年第11届生物医学信号与图像 697
-
2026资源、化学化工与应用材料国际 2559
-
2026年图像处理与数字创意设计国际 2369
-
2026年机械工程,新能源与电气技术 6849
-
2026年材料科学、低碳技术与动力工 2524
-
2026年海洋科学、水利工程与环境管 06-18

-
2026年环境工程、材料科学与循环经 06-18
-
2026年航空动力、流体力学与热物理 06-18
-
2026年地球化学、核物理与地质学国 06-18
-
2026年微机电、物理学与建模仿真国 06-18
-
2026年机械工程、电子技术与自动化 06-18


































-
2026 JCR影响因子正式发布272
-
中国科协发布2025年《重要学术858
-
2026年新锐分区(原中科院期刊5648
-
2025年两院院士增选有效候选人5280
-
好学术:科研网址导航|学术头条分6842
-
2025年国际期刊预警名单发布!7028
-
2025年中科院期刊分区表重磅发24788
-
吉林大学校长张希:学术会议中的提8093
-
研究表明太阳耀斑终端激波可作为地06-24
-
研究揭示藻—菌共生体系强化养殖尾06-24
-
双功能手性双核镍催化研究获进展06-24
-
研究发现银河系中心极端环境下大质06-24
-
废塑料升级利用研究取得进展06-24
-
硒太阳能电池研究取得进展06-24
-
南京大学王涛团队首次发现110亿06-24
-
中华两岸经贸繁荣促进会北京办事处 23333

-
杭州译风会展服务有限公司 18367
-
SHDIF 8410
-
国际工学技术出版协会 24381
-
晔路盛燃气公司 21252
-
北京京西酒店 2626
-
武汉赛思会务有限公司 23739
-
武汉新至恒营销策划有限公司 8322
-
上海脑华商务咨询有限公司 9272
-
中科成创(北京)生物技术有限公司 8297
-
中国医院药学杂志编辑部 23370
-
International As 8325
-
哈尔滨正元会议服务有限责任公司 23583
-
沈阳市东北大学 18557
-
山本五一公司 18380
-
大连全顺会议服务有限公司 23551
-
中国能源学会 24597
-
福州诚信达房产代理有限公司 21396
-
北京师范大学 18442
-
南方电网深圳供电局 2333



