



















北京大学生命科学学院蒋争凡课题组发现相分离启动NLRP3的激活
2025/04/24
 680
680NLRP3炎症小体属于细胞质内的超分子复合物,主要由受体NLRP3蛋白、接头蛋白ASC以及蛋白酶Caspase-1组成。其激活分为两个步骤:首先是启动阶段,Toll样受体(TLR)配体、细胞因子等与相应受体结合,激活NF-κB信号通路,促使NLRP3和IL-1β等蛋白的表达上调,为后续激活奠定物质基础。随后,在多种激活信号的刺激下,NLRP3发生寡聚化,招募ASC和pro-Caspase-1,形成有活性的炎症小体复合物,激活Caspase-1。NLRP3炎症小体作为免疫系统的关键组成部分,在机体抵御病原体入侵和维持内环境稳定方面发挥着不可或缺的作用。NLRP3的异常活化与非常多的炎性疾病,特别是自身免疫病的发生发展直接相关。
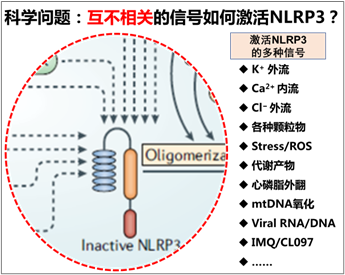
长期以来,尽管针对NLRP3激活机制已开展大量研究,NLRP3炎症小体的激活机制在免疫学领域始终是一个未解之谜,主要原因有以下3个方面:(1)刺激因素的多样性:NLRP3可被众多性质各异的因素激活,涵盖微生物产物、内源性分子和环境刺激物等。在微生物感染相关的病原体相关分子模式(PAMPs)方面,有细菌的脂多糖、肽聚糖、病毒核酸等;内源性的损伤相关分子模式(DAMPs),像细胞内释放的ATP、异常代谢产物如棕榈酸、尿酸晶体等;还有环境刺激物,如紫外线、二氧化硅颗粒等。这些激活物的化学结构和性质差异巨大,比如ATP是一种核苷酸,而二氧化硅是无机物,NLRP3却能对它们都产生反应,这让人们难以确定其识别和响应这些刺激的统一模式。而且,细胞受到这些刺激时会引发多种细胞内变化,这些变化相互交织,使得确定直接导致NLRP3激活的关键因素变得困难。(2)信号通路的复杂性:涉及NLRP3激活的信号通路众多,包括钾离子外流、钙离子信号、线粒体功能障碍、溶酶体破裂、咪喹莫特(IMQ)刺激等,这些通路彼此之间存在复杂的相互作用和重叠。(3)细胞和物种差异的影响:NLRP3的激活机制在不同细胞类型和物种间表现出明显差异。在人类单核细胞中,脂多糖(LPS)刺激可通过释放内源性ATP,激活P2X7受体,从而触发NLRP3炎症小体的激活和IL-1β的成熟;而在小鼠骨髓来源的巨噬细胞中,NLRP3的激活则需要更为严格的两步过程,即先经过TLR等受体的活化进行启动,再由其他激活信号诱导激活。这种差异使得难以归纳出适用于所有情况的统一激活机制,增加了研究的难度。因此,至今尚无一个被广泛认可的统一模型,能够清晰、完整地解释NLRP3如何感知刺激并被激活。不同研究往往侧重不同的信号通路或分子机制,使得该领域的研究较为分散,缺乏一个系统性的理论框架来整合这些研究成果,这也成为深入理解NLRP3激活机制的一大阻碍,限制了人们对炎症相关疾病发病机制的深入理解和有效治疗手段的开发。
2025年4月1日,北京大学生命科学学院教授蒋争凡课题组在Cell Research上以Research Article形式在线发表NLRP3炎症小体领域最新成果“Signal-induced NLRP3 phase separation initiates inflammasome activation”。该论文主要研究NLRP3炎症小体的激活机制,发现信号诱导的NLRP3相分离是其激活的关键,且棕榈酰转移酶ZDHHC7介导的NLRP3棕榈酰化起重要作用,为理解NLRP3激活及相关疾病治疗提供了新方向。
本研究运用全基因组CRISPR-Cas9筛选富集并鉴定棕榈酰转移酶ZDHHC7介导NLRP3炎症小体的激活,去棕榈酰转移酶ABHD13负调控NLRP3炎症小体的激活。通过对人NLRP3蛋白45个半胱氨酸残基及鼠Nlrp3蛋白48个半胱氨酸残基进行突变,利用酰基-生物素交换(ABE)和酰基-聚乙二醇交换(APE)方法,筛选并鉴定出ZDHHC7介导的NLRP3棕榈酰化修饰位点,即人NLRP3的C130和C261和鼠Nlrp3C126(唯一位点)。在ZDHHC7敲除细胞株以及NLRP3-C261S或Nlrp3-C126S突变体中,NLRP3炎症小体的激活均完全受到抑制。含有NLRP3棕榈酰化位点的小分子多肽能够有效抑制NLRP3炎症小体的激活,表明ZDHHC7依赖的棕榈酰化修饰位点是有效的NLRP3小分子抑制剂靶点。此外,他们通过体内实验证实了ZDHHC7对NLRP3激活的重要性,在ZDHHC7基因敲除小鼠模型中,NLRP3炎症小体的激活受到显著抑制。
本研究取得了多项重要发现,首次揭示了信号依赖的NLRP3相分离是其激活的关键,这一过程涉及多个重要分子和事件。棕榈酰转移酶ZDHHC7介导的NLRP3棕榈酰化修饰与NLRP3的FISNA结构域中的一段由12个氨基酸组成的内在无序区域(IDR)在NLRP3相分离和激活过程中发挥着至关重要的作用。其中IDR区域中的三个保守疏水残基对介导多价弱相互作用至关重要,它们的存在促进了NLRP3的相分离,进而推动炎症小体的激活。研究还揭示了多种NLRP3激活刺激物,如钾离子外流、与NLRP3相互作用的分子(咪喹莫特、棕榈酸酯、心磷脂等),均能诱导NLRP3发生构象变化,进而引发相分离和激活。更为重要的是,他们发现两亲性分子,如通常用于抑制生物大分子相分离的1,6-己二醇(1,6-HD)等诸多二醇、化疗药物阿霉素和紫杉醇等可通过降低NLRP3的溶解度,直接诱导其相分离和激活,且这一过程不依赖于ZDHHC7介导的棕榈酰化修饰,与此对应的是,ZDHHC7介导的NLRP3棕榈酰化修饰在静息细胞中持续存在,它通过降低NLRP3相分离的阈值,使NLRP3在受到刺激时更容易被激活。而ABHD13作为一种去棕榈酰化酶,则负向调节NLRP3激活,两者共同维持NLRP3的激活平衡。此外,NLRP3 ATPase酶活特异性抑制剂CY-09无法抑制NLRP3相分离,但可以抑制NLRP3炎症小体激活,表明NLRP3相分离先于其ATP酶活性所引发的ATP/ADP交换及ATP依赖的NLRP3蛋白构象变化,进而招募ASC蛋白并组装成炎症小体复合体。
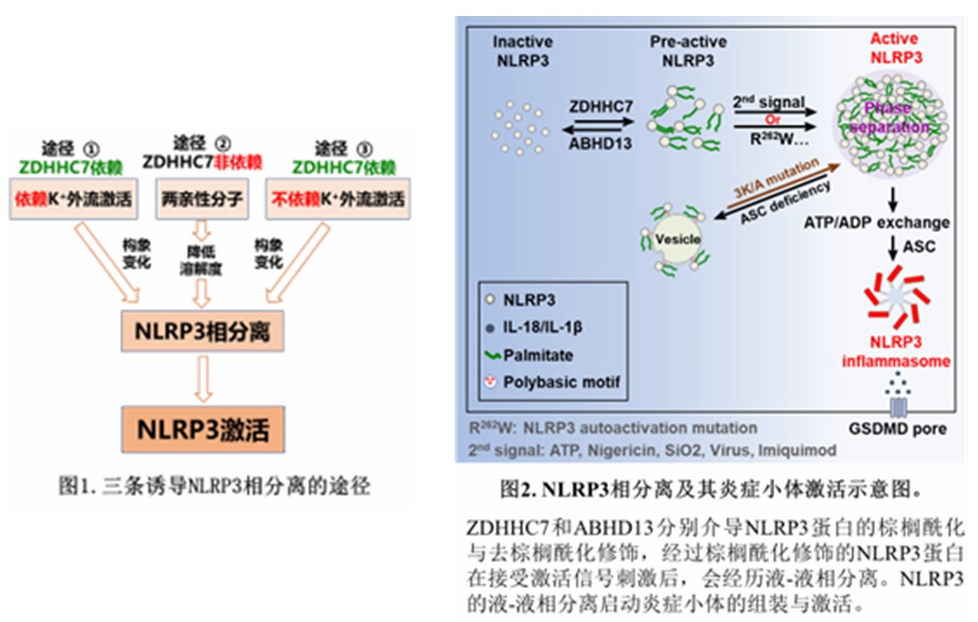
综上所述,该研究展示了3条诱导NLRP3相分离的途径,分别为ZDHHC7依赖且依赖K⁺外流激活、ZDHHC7依赖但不依赖K⁺外流激活以及ZDHHC7非依赖的激活途径(图1)。研究提出了新的统一性NLRP3激活模型(图2),初步回答了“诸多互不相关的刺激如何激活NLRP3”这个重要科学问题。NLRP3相分离激活机制为开发针对炎症相关疾病(如自身免疫性疾病、代谢性疾病和神经退行性疾病等)的新型治疗策略提供了新的理论依据。通过干预NLRP3的相分离过程,有望开发出精准有效的治疗药物,为众多患者带来希望。
北京大学生命科学学院博士后邹恭鲁和2019级博士生唐玉娈为该论文的共同第一作者,蒋争凡为通讯作者。本研究工作得到了科技部国家重点研发计划、国家自然科学基金委、北京大学“细胞增殖与分化”教育部重点实验室及北大-清华生命科学联合中心的资助。
文章来源北京大学,分享只为学术交流,如涉及侵权问题请联系我们,我们将及时修改或删除。







-
2026年第五届机器学习、云计算与智 26

-
2026年第二届计算机视觉与机器学习 627
-
2026年6月优质国际学术会议推荐 1157
-
2026年智慧教育与数据挖掘国际学术 813
-
2026年第11届生物医学信号与图像 697
-
2026资源、化学化工与应用材料国际 2559
-
2026年图像处理与数字创意设计国际 2369
-
2026年机械工程,新能源与电气技术 6849
-
2026年材料科学、低碳技术与动力工 2524
-
2026年海洋科学、水利工程与环境管 06-18

-
2026年环境工程、材料科学与循环经 06-18
-
2026年航空动力、流体力学与热物理 06-18
-
2026年地球化学、核物理与地质学国 06-18
-
2026年微机电、物理学与建模仿真国 06-18
-
2026年机械工程、电子技术与自动化 06-18


































-
2026 JCR影响因子正式发布272
-
中国科协发布2025年《重要学术858
-
2026年新锐分区(原中科院期刊5648
-
2025年两院院士增选有效候选人5280
-
好学术:科研网址导航|学术头条分6842
-
2025年国际期刊预警名单发布!7028
-
2025年中科院期刊分区表重磅发24788
-
吉林大学校长张希:学术会议中的提8093
-
研究表明太阳耀斑终端激波可作为地06-24
-
研究揭示藻—菌共生体系强化养殖尾06-24
-
双功能手性双核镍催化研究获进展06-24
-
研究发现银河系中心极端环境下大质06-24
-
废塑料升级利用研究取得进展06-24
-
硒太阳能电池研究取得进展06-24
-
南京大学王涛团队首次发现110亿06-24
-
北京嘉园文华发展中心 2387

-
烟台群英会议服务有限公司 24263
-
北京清华大学计算机系 21489
-
河北百信米斯文化传播有限公司 8432
-
美通社北京分公司 2401
-
天津中豪会议策划服务有限公司 2790
-
北京北发电子商务股份有限公司 18396
-
csae@ieee-csae.o 2492
-
先进科学与技术研究中心 24451
-
新电子杂志 18591
-
华北电力大学 21471
-
21255
-
中华医学会行为医学分会 18510
-
中国广东东莞 18445
-
首都师范大学教育科学学院 21523
-
沈阳博思教育咨询有限公司 23306
-
中华人物组委会 18295
-
宁夏大学人文学院 23434
-
北京市小汤山特菜大观园 21528
-
云南财经大学 21484



